
Александр Марков
Ангедония — утрата способности получать удовольствие — считается одним из главных симптомов депрессии. Давно известно, что ангедония часто развивается после стресса, однако конкретные популяции нейронов, ответственные за этот эффект, долго не удавалось выявить.
Главными «подозреваемыми» были дофаминэргические нейроны вентральной области покрышки среднего мозга (ventral tegmental area, VTA), однако эксперименты не давали этому однозначного подтверждения. Американские нейробиологи показали в опытах на мышах, что за послестрессовую ангедонию отвечают не дофаминэргические, а ГАМК-эргические нейроны VTA, посылающие тормозные сигналы в прилежащее ядро (nucleus accumbens, NAc). Если подавить активность этих нейронов во время стресса, стресс не вызовет ангедонии, а если искусственно стимулировать их с определенной частотой (4 Гц), ангедония возникает даже без стресса. Ранее про эти нейроны было известно, что они возбуждаются в ответ на неприятные стимулы, задействованы в формировании зависимостей, а также отвечают за уменьшение удовольствия от ожидаемой награды по сравнению с нежданными подарками судьбы.

Частичная или полная утрата способности получать удовольствие от чего бы то ни было (ангедония) — один из самых неприятных симптомов депрессии и некоторых других психических расстройств. Нейробиологические механизмы ангедонии остаются неясными, хотя в целом понятно, что дело скорее всего в каких-то нарушениях в работе системы внутреннего подкрепления (см. Reward system).
Вряд ли это утешит людей в депрессии, но нейробиологию ангедонии вполне можно изучать на животных моделях, таких как мыши. Хоть мышь и не скажет экспериментатору, что жизнь ей больше не мила, об этом можно судить по подавленности поиска награды (reward-seeking behaviour), то есть поведения, направленного на получение удовольствия.
У грызунов, как и у людей, острый стресс подавляет поиск награды ненадолго, а хронический может вызвать длительную ангедонию. Многие ученые пытались расшифровать нейробиологические основы этих эффектов. Ключевую роль в поиске награды, мотивации поведения, обучении на положительном опыте и формировании зависимостей играют дофаминовые сигналы, посылаемые дофаминэргическими нейронами вентральной области покрышки (Ventral tegmental area, VTA) в прилежащее ядро (Nucleus accumbens, NAc) и некоторые другие отделы мозга. Поэтому логично предположить, что стресс приводит к ангедонии, влияя на работу именно этих нейронов (R. A. Wise, 2008. Dopamine and reward: the anhedonia hypothesis 30 years on). Однако многолетние исследования в этом направлении так и не дали однозначных результатов.
В новом исследовании, результаты которого опубликованы 10 июня в журнале Nature Communications, американские нейробиологи решили не ограничиваться регистрацией активности дофаминовых нейронов VTA, а провести более широкий скрининг процессов, происходящих при стрессе в разных отделах мозга, так или иначе связаных с депрессией: в префронтальной коре, NAc, дорзальном и вентральном гиппокампе, базолатеральной миндалине и VTA. Вставленные в мозг мышей электроды регистрировали как активность отдельных нейронов, так и внеклеточные «потенциалы ближнего поля» (Local field potential, LFP; этот метод еще называют «внутричерепной электроэнцефалографией»), которые отражают состояние больших популяций нейронов, расположенных по соседству с электродом (см. G. Buzsáki et al., 2012. The origin of extracellular fields and currents — EEG, ECoG, LFP and spikes). В частности, по низкочастотным колебаниям LFP можно судить о синхронном поступлении входных сигналов (то есть тех или иных нейромедиаторов) в изучаемый участок мозга.
Подопытные мыши при этом либо находились в комфортной обстановке, либо подвергались острому стрессу: несчастных запихивали в тесную камеру, где они едва могли пошевелиться, или подвешивали за хвост на 30 минут. Эти не очень гуманные эксперименты (одобренные, впрочем, компетентными органами в соответствии с требованиями NIH) показали, что острый стресс приводит к сильным колебаниям LFP в прилежащем ядре с частотой около 4 Гц (рис. 2, вверху). В некоторых других отделах мозга тоже были замечены похожие колебания, но в NAc они выражены намного сильнее. Колебания затухают, когда мышь дергается, пытаясь высвободиться, и усиливаются, когда животное неподвижно. Всё это наблюдалось как у мышей, запертых в тесной камере, так и у подвешенных за хвост. В комфортной обстановке этих колебаний нет ни у двигающихся, ни у неподвижных мышей. Таким образом, четырехгерцовые колебания LFP в NAc, по-видимому, отражают именно реакцию на острый стресс.
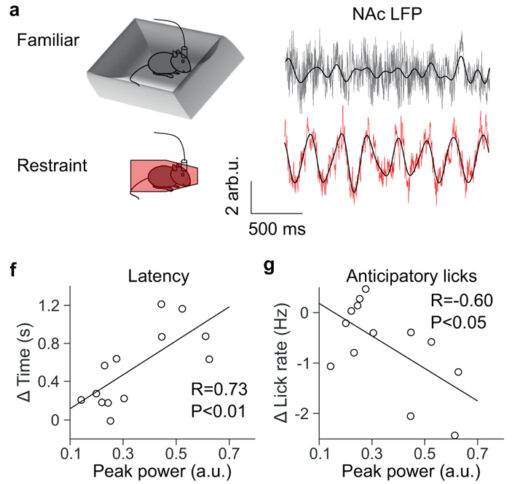
Ученые решили проверить, нет ли связи между колебаниями LFP и ангедонией. Для этого мышей приучили к тому, что они могут получить награду (каплю воды) после звукового сигнала, если вовремя лизнут сухую поилку. Чтобы усилить мотивацию, мышам перед обучением, конечно, не давали пить вволю. Награду можно было получить в течение пяти секунд, в интервале от 1,5 до 6,5 секунд после включения звукового сигнала. Специальный датчик регистрировал прикосновение мышиного язычка и выдавал 15 мкл воды.
Как и ожидалось, у мышей, только что переживших стресс, поиск награды был подавлен: они позже начинали лизать поилку и совершали меньше «упреждающих лизаний» до появления награды (хотя общая подвижность у них не снижалась). Главный новый результат состоял в том, что обнаружилась отрицательная корреляция между силой колебаний LFP в NAc во время стресса и интенсивностью поиска награды впоследствии. Чем сильнее были колебания LFP, тем ярче были выражены симптомы ангедонии (рис. 2, внизу). Иными словами, по силе колебаний LFP во время стресса можно предсказать, в какой степени будет подавлен поиск награды после стресса. Эффект исчезал на следующий день, то есть ангедония была непродолжительной, как и полагается после кратковременного стресса.
Выяснилось также, что активность (частота потенциалов действия, firing rate) большинства (59%) нейронов NAc снижается во время стресса и при этом колеблется в точном соответствии с колебаниями LFP. У 21% нейронов NAc активность, наоборот, повышается при стрессе, а у 20% она не меняется. Но у этих двух категорий нейронов активность не коррелирует с колебаниями LFP, а значит, вряд ли напрямую связана с ангедонией.
Эти данные позволяют предположить, что четырехгерцовые колебания LFP отражают ритмическое поступление в NAc тормозных сигналов. А важнейшим тормозным медиатором в центральной нервной системе, как известно, является ГАМК.
Дальнейшие эксперименты подтвердили эту гипотезу. Более того, они выявили источник тормозных сингалов. Им оказались ГАМК-эргические нейроны VTA — крайне интересная, хотя и недостаточно изученная популяция нервных клеток. Известно, что эти нейроны «выстреливают» в ответ на неприятные стимулы. Есть серьезные подозрения, что они вовлечены в формирование зависимостей под действием стресса. Кроме того, они возбуждаются в ответ на условные сигналы, предвещающие награду. Именно они виновны в уменьшении чувства удовольствия от награды в том случае, если животное заранее уверено в ее появлении (см. В «системе вознаграждения» найдены нейроны, возбуждающиеся от хороших предчувствий, «Элементы», 10.02.2012). И вот теперь на них возложили ответственность еще и за послестрессовую ангедонию.
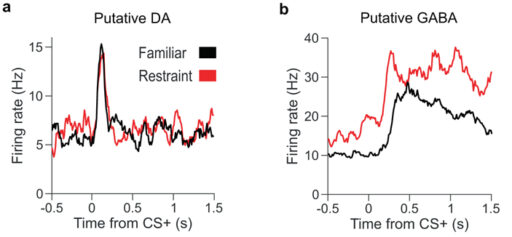
Этот вывод был подтвержден множеством разнообразных экспериментов, в том числе с использованием методов оптогенетики. Например, исследователи неизбирательно подавляли работу всех нейронов VTA при помощи инъекции мусцимола — и это приводило к уменьшению амплитуды четырехгерцовых колебаний LFP в NAc при стрессе. Но если подавить только работу дофаминовых нейронов VTA (65% нейронов VTA — дофаминэргические), то вызываемые стрессом колебания LFP в NAc не уменьшаются. Активность дофаминовых нейронов VTA в целом снижается при стрессе, однако она не коррелирует с колебаниями LFP в NAc. Напротив, колебания активности ГАМК-эргических нейронов VTA (они составляют 25% от общего числа нейронов VTA, и аксоны многих из них идут в NAc) коррелируют с колебаниями LFP. Если избирательно заблокировать работу этих нейронов во время стресса, то колебания LFP резко уменьшаются. Самое интересное, что после такого стресса, при котором работа ГАМК-эргических нейронов VTA была искусствено подавлена, поиск награды у мышей не только не ослабевает, но даже усиливается по сравнению с контролем.
Эти эксперименты показали, что работа ГАМК-эргических нейронов VTA необходима для развития послестрессовой ангедонии. Были поставлены также опыты, показавшие, что она для этого достаточна. Оказалось, что если искусственно возбуждать ГАМК-эргические нейроны VTA c частотой 4 Гц, то это, во-первых, создает характерные колебания LFP в NAc, во-вторых — подавляет поиск награды, то есть вызывает ангедонию, причем без всякого стресса. Ученые испытали также частоту 20 Гц, но это не сработало: колебания LFP в NAc не появились, а поиск награды уменьшился едва заметно. По-видимому, здесь важно не просто поступление ГАМК в NAc, а его ритмичность. Это, кстати, объясняет, почему в прежних исследованиях не удавалось обнаружить связь между искусственным возбуждением ГАМК-эргических нейронов VTA и поиском награды: экспериментаторы просто возбуждали нейроны не в том ритме.
Дофаминэргические нейроны VTA реагируют на условный сигнал, предвещающий награду, лишь кратковременным всплеском активности, а у ГАМК-эргических повышенная активность сохраняется в течение всего периода ожидания награды (об исследовании, в котором это было показано, рассказывает новость В «системе вознаграждения» найдены нейроны, возбуждающиеся от хороших предчувствий, «Элементы», 10.02.2012). Как говорят нейробиологи, активность ГАМК-эргических нейронов VTA «кодирует ожидание награды». Именно эта активность в итоге делает ожидаемую награду менее приятной, чем нежданные подарки судьбы.
Всё это подтвердилось в обсуждаемой работе. Но авторы продвинулись дальше, связав реакцию ГАМК-эргических нейронов VTA на предвещающий награду стимул с послестрессовой ангедонией. Авторы предположили, что «кодирование ожидания награды» ГАМК-эргическими нейронами VTA меняется после стресса. Это подтвердилось. Выяснилось, что после пережитого стресса реакция ГАМК-эргических нейронов VTA на условный сигнал, предвещающий награду, значимо усиливается. Что касается дофаминэргических нейронов, то их реакция на такой сигнал не меняется после стресса (рис. 3).
Во всем этом можно усмотреть некую логику, связывающую воедино две функции ГАМК-эргических нейронов VTA: уменьшение радости от ожидаемой награды и послестрессовую ангедонию. Получается, что, с точки зрения этих нейронов, после острого стресса ожидаемая награда становится как бы еще более ожидаемой. А значит — скучной и не сулящей радости. Возможно, читателям, страдающим депрессией, это покажется знакомым.
В обсуждаемой работе рассматривается только непродолжительный стресс и вызываемое им кратковременное угнетение поиска награды. Хронический стресс, который у мышей, как и у людей, может вызвать стойкую ангедонию, в работе не изучался. Очень может быть, что ГАМК-эргические нейроны VTA играют важную роль и в этом случае, но для проверки понадобятся дополнительные исследования.
По прочтении этой заметки кому-то из читателей, возможно, захочется поискать в литературе данные об ингибиторах ГАМК-эргических нейронов VTA, которые могли бы сделать жизнь более радостной. Но это чревато разочарованием, потому что такие ингибиторы вообще-то человечеству давно известны, и ничего хорошего их применение не сулит (S. H. Stobbs et al., 2004. Ethanol suppression of ventral tegmental area GABA neuron electrical transmission involves N-methyl-D-aspartate receptors; S. C. Steffensen et al., 2006. Contingent and non-contingent effects of heroin on mu-opioid receptor-containing ventral tegmental area GABA neurons).
Источник: Daniel C. Lowes, Linda A. Chamberlin, Lisa N. Kretsge, Emma S. Holt, Atheir I. Abbas, Alan J. Park, Lyubov Yusufova, Zachary H. Bretton, Ayesha Firdous, Armen G. Enikolopov, Joshua A. Gordon & Alexander Z. Harris. Ventral tegmental area GABA neurons mediate stress-induced blunted reward-seeking in mice // Nature Communications. 2021. V. 12. Article number: 3539
Источник: Элементы https://elementy.ru/novosti_nauki/433827/Naydeny_neyrony_otvetstvennye_za_poslestressovuyu_angedoniyu.